Impact de Mycoplasma ovipneumoniae sur la survie des juvéniles du mouflon d’Amérique (Ovis canadensis) dans l’écosystème du bassin et de l’aire de répartition nord [PeerJ]
![Impact de Mycoplasma ovipneumoniae sur la survie des juvéniles du mouflon d’Amérique (Ovis canadensis) dans l’écosystème du bassin et de l’aire de répartition nord [PeerJ]](https://guidesurvie.com/wp-content/uploads/2021/02/Impact-de-Mycoplasma-ovipneumoniae-sur-la-survie-des-juveniles-du.jpg)
introduction
L’étude de la dynamique des populations est essentielle pour la gestion des espèces (Williams, Nichols et Conroy, 2002). Le recrutement est un processus crucial pour la dynamique des populations, par lequel les populations gagnent des individus grâce aux naissances et à l’immigration (Pradel, 1996). Le recrutement varie fortement selon les espèces selon qu’elles sont sélectionnées k, c’est-à-dire avoir moins de jeunes avec un plus grand investissement parental, ou r-sélectionnées, c’est-à-dire avoir plus de jeunes avec un investissement parental réduit (MacArthur et Wilson, 1967). Pour les espèces sélectionnées par k, comme les grands herbivores terrestres, la survie annuelle des adultes a tendance à être relativement élevée avec peu de variation. En revanche, la survie des juvéniles a tendance à être plus variable et, par conséquent, la croissance de la population a tendance à être plus sensible aux paramètres de survie des juvéniles (Gaillard et al., 2000). Par conséquent, il est essentiel de prendre en compte les variables affectant la survie des juvéniles lors de la gestion de ces espèces.
Mouflon d’Amérique (Ovis canadensis) ont un rendement reproductif relativement faible (Festa-Bianchet et al., 2019). Les femelles ont rarement plus d’une progéniture par an et peuvent ne pas atteindre leur plein potentiel de reproduction avant leur 4e année (Rubin, Boyce et Bleich, 2000). Ainsi, la survie des juvéniles peut avoir un impact significatif sur les trajectoires des populations (Manlove et coll., 2019). La survie des juvéniles est influencée par la maladie (Cassirer et Sinclair, 2007; Smith et coll., 2014), condition corporelle maternelle (Festa-Bianchet, 1998), qualité du fourrage (Feder et coll., 2008), conditions météorologiques (Douglas, 2001), diversité génétique (Hogg et coll., 2006) et la prédation (Rominger, 2018). La maladie, en particulier la pneumonie, peut avoir un effet dramatique sur la survie des juvéniles (Manlove et coll., 2016; Garwood et coll., 2020). Plus généralement, les maladies respiratoires ont probablement causé le déclin du mouflon d’Amérique dans l’ouest de l’Amérique du Nord et continuent d’entraver le rétablissement de l’espèce (Cassirer et coll., 2018). Par conséquent, l’évaluation de l’interaction de la maladie et d’autres facteurs agissant simultanément sur la survie des juvéniles est essentielle pour la conservation et la gestion des mouflons d’Amérique.
Mycoplasma ovipneumoniae, un pathogène bactérien, est considéré comme le principal agent causal de la pneumonie respiratoire chez le mouflon d’Amérique (Besser et al., 2008; Cassirer et coll., 2018). Transmission de M. ovipneumoniae de moutons domestiques (O. bélier) et les chèvres (Capra hircus) aux mouflons d’Amérique est généralement suivie d’une mortalité élevée d’individus de toutes les classes d’âge dans les populations non immunisées. Certains survivants élimineront la maladie, tandis que d’autres restent des porteurs chroniques qui continuent à excréter l’agent pathogène même s’ils semblent souvent relativement sains (Besser et al., 2013). Les porteurs chroniques peuvent ainsi transmettre M. ovipneumoniae et maintenir sa présence au sein d’une population, en particulier par transmission à des juvéniles auparavant non exposés (Plowright et al., 2013). Ce schéma d’infection chez le mouflon d’Amérique a été observé dans plusieurs régions, notamment le nord-est de l’Oregon, l’Idaho, l’État de Washington, le Nevada et le Dakota du Sud (Cassirer et coll., 2018; Garwood et coll., 2020). Immunité à M. ovipneumoniae est considérée comme spécifique à la souche, de nouvelles souches entraînant de nouvelles vagues d’infection (Cassirer et coll., 2017).
Réponses au niveau de la population M. ovipneumoniae les flambées, cependant, peuvent varier considérablement. Des variations considérables ont été observées dans les niveaux de mortalité tous âges au premier contact, la survie ultérieure des adultes et la survie des juvéniles au cours des années suivantes parmi les populations et entre les lignées évolutives et les habitats (Cassirer et coll., 2018; Dekelaita et al., 2020). Cette variation a été supposée provenir de nombreuses causes, y compris la virulence de la souche (Kamath et coll., 2019), des facteurs nutritionnels tels que la qualité du fourrage et la densité de la population (Dekelaita et al., 2020), des facteurs stochastiques tels que la présence de porteurs chroniques (Cassirer et coll., 2018; Garwood et coll., 2020), diversité génétique des populations hôtes (Cassirer et coll., 2018) et des différences phénologiques entraînant différents modèles d’agrégation, de contact et de dispersion (Cassirer et coll., 2018). En effet, les mouflons d’Amérique habitent des écosystèmes allant des déserts arides du nord du Mexique et du sud-ouest des États-Unis d’Amérique aux glaciales montagnes Rocheuses du nord de l’Alberta et présentent une variation phénotypique significative et des preuves d’adaptation locale (Wehausen et Ramey II, 2000; Wiedmann et Sargeant, 2014; Malaney et coll., 2015). Actuellement, trois sous-espèces de mouflons d’Amérique sont reconnues, le mouflon du désert (O. c. Nelsoni), le mouflon d’Amérique des Rocheuses (O. c. canadensis) et le mouflon d’Amérique de la Sierra Nevada (O. c. Sierrae) (Wehausen et Ramey II, 2000; Wehausen, Bleich et Ramey II, 2005). Auparavant, d’autres sous-espèces étaient reconnues (par exemple, le mouflon d’Amérique de la péninsule, O. c. cremnobates, et mouflons d’Amérique de Californie, O. c. Californie) (Cowan, 1940). Le débat reste à savoir si ces lignées putatives reflètent d’importantes trajectoires évolutives indépendantes et une adaptation locale importante (Buchalski et coll., 2016; Bleich, Sargeant et Wiedmann, 2018; Barbosa et al., Sous presse).
Études antérieures de la dynamique des populations en présence de M. ovipneumoniae suggèrent que la dynamique des maladies devrait être évaluée à travers les lignées et les écosystèmes. Par exemple, le mouflon d’Amérique des montagnes Rocheuses du système Hells Canyon de l’Idaho, de l’Oregon et de Washington occupe un habitat relativement continu avec des hivers froids (Cassirer et Sinclair, 2007). Dans ce système, M. ovipneumoniae a tendance à persister pendant de longues périodes, entraînant une maladie constante chez les juvéniles (Cassirer et coll., 2018). Le même schéma d’infection a également été observé chez le mouflon d’Amérique des Rocheuses dans les Black Hills de l’ouest du Dakota du Sud et de l’est du Wyoming (Smith et coll., 2014; Garwood et coll., 2020). Dans le désert de Mojave, dans le sud-ouest des États-Unis d’Amérique, les mouflons d’Amérique occupent des poches de chaînes de montagnes isolées entourées d’un désert de basse altitude mais liées par des mouvements inter-montagnes, ce qui entraîne des métapopulations naturelles (Bleich et coll., 1996). M. ovipneumoniae dans ce système semble avoir été présent périodiquement (Shirkley et al., Sous presse), et les impacts sur la survie des adultes et le recrutement des juvéniles semblent être très variables (Dekelaita et al., 2020).
Les mouflons d’Amérique du nord de l’écosystème du bassin et de la chaîne de montagnes (y compris des parties du sud-est de l’Oregon, du sud-ouest de l’Idaho et du nord du Nevada) occupent des habitats de transition entre les systèmes Mojave et Rocky Mountain et ont une histoire unique. Le mouflon d’Amérique de cette région était considéré comme la sous-espèce «Californie» (O. c. Californie; Cowan, 1940), s’étendait de la Sierra Nevada de Californie au nord jusqu’à la Colombie-Britannique. En Oregon, tous les mouflons d’Amérique ont disparu en 1945 (Département de la pêche et de la faune de l’Oregon, 2003). Par la suite, l’analyse morphométrique et ADN des populations indigènes éteintes de l’Oregon a abouti à la réaffectation de ces populations à la forme désertique du Grand Bassin du mouflon d’Amérique (O. c. Nelson; Wehausen et Ramey II, 2000). Les mouflons d’Amérique qui existent maintenant dans l’Oregon sont tous issus de translocations et sont gérés comme deux lignées: les mouflons d’Amérique des Rocheuses ont été introduits dans le nord-est de l’Oregon et les mouflons d’Amérique de Californie ont été introduits dans l’habitat potentiel ou ancien du mouflon d’Amérique dans d’autres régions de l’État à l’aide d’individus. de la Colombie-Britannique. À l’heure actuelle, les deux lignées sont officiellement considérées comme des sous-espèces de mouflons d’Amérique des Rocheuses (O. c. canadensis; Wehausen et Ramey II, 2000). La plupart des populations rétablies de l’Oregon provenaient d’une seule translocation de 22 mouflons d’Amérique de la Colombie-Britannique en 1953 (Olson, Whittaker et Rhodes, 2013). Cependant, en 2000 et 2001, dans le cadre d’un effort expérimental visant à accroître la diversité génétique de la population et à améliorer la performance démographique, deux populations du sud-est de l’Oregon ont reçu des augmentations de mouflons d’Amérique transférés provenant de différentes populations sources de la Colombie-Britannique (Olson, Whittaker et Rhodes, 2012). Des mouflons d’Amérique ont également été introduits dans le nord du Nevada en 1972 à partir de plusieurs populations sources (NDOW, 2001; Olson, Whittaker et Rhodes, 2013) et une dispersion de ces populations dans le sud-est de l’Oregon a été observée (ODFW, données non publiées). Les populations de mouflons d’Amérique du sud-ouest de l’Idaho et du nord du Nevada partagent des histoires similaires, bien que les translocations vers le nord du Nevada reposent sur un plus grand nombre de populations sources que l’Oregon.
L’habitat du mouflon d’Amérique dans l’écosystème du nord du bassin et de l’aire de répartition présente une structure semblable à une métapopulation, où le mouflon d’Amérique utilise des parcelles discrètes de terrain d’évacuation escarpé séparées par de vastes zones de prairies ou d’armoise. En raison de leur histoire démographique et de leur répartition spatiale, les mouflons d’Amérique du sud-est de l’Oregon et du nord du Nevada ont une faible diversité génétique (Olson, Whittaker et Rhodes, 2013; Malaney et coll., 2015) par rapport aux niveaux mesurés dans les études sur le mouflon d’Amérique des Rocheuses dans les montagnes Rocheuses du Colorado (Driscoll et coll., 2015) et le mouflon d’Amérique du désert dans le désert de Mojave en Californie (Epps, Crowhurst et Nickerson, 2018). Les mouflons d’Amérique dans l’écosystème nordique du bassin et de l’aire de répartition connaissent également une phénologie des plantes fourragères différente de celle observée dans les systèmes du désert ou des montagnes Rocheuses. On peut donc s’attendre à ce que la dynamique de la maladie diffère également.
Bien que la mortalité par cause chez les juvéniles de mouflons d’Amérique ait été largement étudiée dans d’autres systèmes (Smith et coll., 2014; Cassirer et coll., 2018; Cain et coll., 2019; Garwood et coll., 2020), il n’a pas été évalué chez des mouflons d’Amérique élevés en Californie. Dans cette étude, nous évaluons la distribution et l’influence de M. ovipneumoniae sur 13 populations de mouflons d’Amérique gérées par la «Californie» dans le sud-est de l’Oregon et le nord du Nevada, et enquêter sur la mortalité des juvéniles en relation avec la qualification du fourrage et la diversité génétique. Bien que la présence de M. ovipneumoniae avait déjà été vérifié dans au moins une population, l’effet des maladies respiratoires sur le système était inconnu. De même, la diversité génétique des populations de ce système était inconnue, même si on s’attendait à ce qu’elle soit faible, et la qualité du fourrage était censée varier considérablement étant donné la large gamme d’altitude utilisée par les mouflons d’Amérique et la forte influence des précipitations dans ce système semi-aride. Nous avons utilisé la télémétrie et des observations sur le terrain pour surveiller les juvéniles de femelles adultes munies d’un collier GPS afin d’estimer la probabilité de survie bimensuelle sur une période de 3 ans. Nous avons émis l’hypothèse que la survie des juvéniles serait influencée par la maladie, la nutrition et la diversité génétique. Nous avons prédit que la probabilité de survie des juvéniles à 4 mois serait plus faible dans les populations qui (1) étaient exposées à M. ovipneumoniae, (2) avaient une hétérozygotie attendue plus faible, et (3) une qualité de fourrage inférieure, comme l’indique l’indice de végétation différentielle normalisé (NDVI) avant et après la mise bas. De plus, après avoir observé la mortalité d’une femelle adulte suspectée d’être une porteuse chronique dans l’une des populations de l’étude à la fin de la période de 3 ans, nous avons mené une enquête de suivi limitée sur la survie des juvéniles dans cette population l’année suivante. .
Matériaux et méthodes
Zone d’étude
Les populations de mouflons d’Amérique que nous avons étudiées étaient situées dans le sud-est de l’Oregon et le nord du Nevada, entre 41,3 et 42,8 ° N, et 117,0 et 118,2 ° W (figure 1). L’ensemble de la zone d’étude faisait partie du bassin et de l’aire de répartition nord (classification de niveau III des écorégions dans Omernik et Griffith, 2014). Cinq populations de notre étude se situaient dans ou en grande partie dans le haut plateau de lave disséqué (Blue Mountain (BSP), Bowden Hills (BHP), Rattlesnake (RSP), Three Forks (TFK), Upper Owyhee (UOP), Fig.1), bien que le BSP et le RSP se soient en partie produits dans l’écorégion des hautes plaines de lave (Omernik et Griffith, 2014). La population de Ten Mile (TMP) faisait partie des hautes plaines de lave, tandis que les hautes plaines de lave et les hautes terres semi-arides (Omernik et Griffith, 2014) dominent l’habitat du mouflon d’Amérique dans le ruisseau Trout (Trout Creek – est (TCE), – sud (TCS) et – ouest (TCW), Fig.1) et la métapopulation de Santa Rosa (Calicos (CAL), Eight Mile (EML), Martin Creek (MCK) et Sawtooth (SAW), Fig.1).
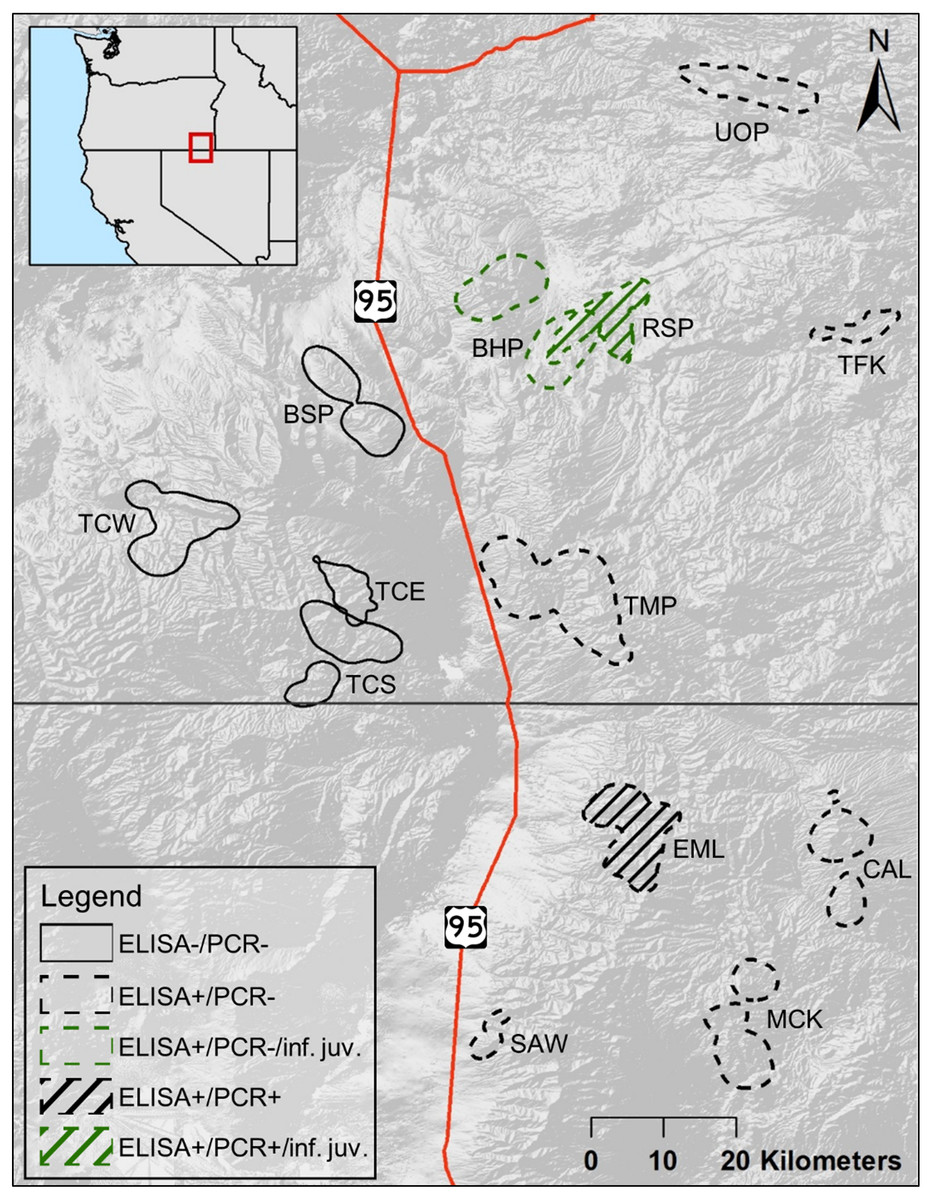
Figure 1: Distributions d’utilisation estivale cumulative des populations de mouflons d’Amérique femelles adultes considérées dans cette étude.
Les polygones qui se chevauchent indiquent un habitat partagé. Des polygones pleins, tous à l’ouest de la route 95 des États-Unis, indiquent des populations non exposées à Mycoplasma ovipneumoniae, avec des lignes pointillées indiquant les populations exposées à l’est de la route 95 des États-Unis. Le remplissage des lignes indique les populations, EML et RSP, où une seule femelle adulte M. ovipneumoniae une infection a été détectée. Les polygones de couleur verte indiquent les populations, BHP et RSP, où les juvéniles morts infectés par M. ovipneumoniae ont été détectés. Les populations comprennent: Bowden Hills (BHP), Blue Mountain (BSP), Calicos (CAL), Eight Mile (EML), Martin Creek (MCK), Rattlesnake (RSP), Sawtooth (SAW), Trout Creeks-east (TCE), Trout Creeks – sud (TCS), Trout Creeks – ouest (TCW), Three Forks (TFK), Ten Mile (TMP) et Upper Owyhee (UOP).
Les écorégions du haut plateau de lave disséqué et des hautes plaines de lave sont toutes deux caractérisées par des plateaux élevés, mais le haut plateau de lave disséqué contient des canyons à parois abruptes ainsi que des lacs intermittents, tandis que les hautes plaines de lave contiennent des cônes et des buttes volcaniques isolés ainsi que des lacs intermittents. et des flux éphémères (Omernik et Griffith, 2014). Des montagnes d’altitude basse à moyenne, généralement avec des pentes raides, et certains cours d’eau éphémères et pérennes caractérisent les terres semi-arides (Omernik et Griffith, 2014). Les types de géologie les plus courants dans les trois écotypes sont le basalte et la rhyolite, entrecoupés d’autres types de roches. Les sols dérivés de ces types de roches sont assez peu profonds et pauvres (Omernik et Griffith, 2014). Les précipitations moyennes dans la zone d’étude sont généralement de 22,5 à 35,0 cm par an, bien que certaines zones des ruisseaux Trout et des montagnes Santa Rosa reçoivent beaucoup plus de précipitations (Omernik et Griffith, 2014).
Les trois écotypes contiennent une steppe d’armoise. Grande armoise (Artemisia tridentata) et d’armoise basse (A. arbusculaire) sont les espèces herbacées ligneuses les plus courantes, tandis que les espèces herbacées indigènes communes sont principalement constituées de graminées vivaces au goût agréable telles que la fétuque de l’Idaho (Festuca idahoensis), agropyre (Psudoroegneria spicata), queue d’écureuil (Elymus Elymoides), Thurber aiguilletée (Achnatherum thurberianum) et le pâturin Sandberg (Poa secunda). Genévrier de l’Ouest (Juniperus occidentalis) est la plante ligneuse la plus commune dans 3 écotypes, que l’on trouve généralement dans les zones rocheuses, tandis que les hautes terres semi-arides se distinguent par des saules (Salix spp.) dans les zones riveraines et le peuplier faux-tremble (Populus tremuloides) et l’acajou de montagne (Cercocarpus spp.) dans les poches de neige (Omernik et Griffith, 2014). Les espèces d’ongulés présentes dans la zone d’étude comprennent le wapiti (Cervus canadensis), cerf mulet (Odocoileus hemionus) et pronghorn (Antilocapra americana) (Omernik et Griffith, 2014). Les prédateurs potentiels du mouflon d’Amérique dans la zone d’étude comprennent le couguar (Puma concolor), les coyotes (Canis latrans), lynx roux (Lynx rufus) et les aigles royaux (Aquila chrysaetos; Omernik et Griffith, 2014).
Les pratiques d’utilisation des terres les plus courantes sont l’élevage de bétail et la culture de céréales et de foin (Omernik et Griffith, 2014). Le pâturage intensif de ces terres et la suppression des incendies naturels ont conduit à la propagation de grands incendies incontrôlables et à l’empiètement par des plantes annuelles envahissantes, telles que la cheatgrass (Bromus tectorum) et medusahead (Taeniatherium caput-medusae; Omernik et Griffith, 2014). Ces graminées surpassent la végétation indigène après un incendie, ce qui conduit à la domination de ces graminées. La faune et le bétail ici dépendent des sources, des zones humides et des sources d’eau artificielles (Omernik et Griffith, 2014).
Historique des populations de mouflons d’Amérique dans la zone d’étude
Le rétablissement des mouflons d’Amérique dans la zone d’étude a commencé en 1978 avec le transfert des mouflons d’Amérique dans la région de Eight Mile de Santa Rosas (Fig. 1). Les trois populations de Trout Creek, TCE, TCS et TCW, et RSP ont été établies avec des translocations uniques chacune de Hart Mountain (tableau S1) en 1987 et 1992. La population de mouflons d’Amérique de Hart Mountain a été établie en 1954 avec une translocation de mouflons d’Amérique de Williams Lake, Colombie-Britannique. Bien que les mouflons d’Amérique du TMP et de l’UOP aient été transférés de diverses autres populations de l’Oregon, toutes ces populations ont finalement été dérivées de la population établie à Hart Mountain en 1954. Les mouflons d’Amérique de la Californie, bien que transférés de la pinède du Nevada, sont également dérivés de Hart Mountain. Les populations de mouflons d’Amérique restantes dans l’étude ont été établies à partir d’une source originale différente, par exemple, MCK et SAW, qui avaient des stocks de Kamloops, en Colombie-Britannique, et de Penticton, en Colombie-Britannique, respectivement, ou de plus d’une source, par exemple, EML. Deux populations ont été établies par dispersion dans un habitat inoccupé: le BSP a été établi avec des événements de dispersion présumés des populations de Trout Creek entre le milieu et la fin des années 1990 (comm. Pers. S. Torland, ODFW), et le BHP aurait été établi par dispersion. mouflons d’Amérique de RSP. D’autres mouvements observés comprennent la dispersion des mâles adultes à collier entre la chaîne de montagnes Santa Rosa et les chaînes de montagnes voisines dans le sud-est de l’Oregon, observée en 2009 et 2010 (NDOW, données non publiées).
Capture et échantillonnage
Tous les tests de capture, de manipulation et de maladie ont été réalisés par le Département des pêches et de la faune de l’Oregon (ODFW) et le Département de la faune du Nevada (NDOW). La méthodologie de capture a suivi les recommandations de Foster (2004) et l’American Society of Mammalogists (Sikes et le Comité de protection et d’utilisation des animaux de l’American Society of Mammalogists, 2016). ODFW et NDOW ont capturé, porté un collier et échantillonné des mouflons d’Amérique femelles adultes dans 13 populations du sud-est de l’Oregon et du nord du Nevada entre janvier 2016 et février 2018 (figure 1). Les captures ont été effectuées à l’aide d’un fusil à filet tiré d’un hélicoptère, avec des mouflons d’Amérique les yeux bandés et entravés une fois capturés (Krausman, Hervert et Ordway, 1985). Les bighorns ont été amenés dans une zone centralisée à la base de leur aire de répartition pour être équipés d’un collier de télémétrie et pour collecter des échantillons biologiques, sauf lorsque l’emplacement de capture était trop éloigné du camp de base pour les transporter rapidement, auquel cas ils ont été traités sur le terrain, à la emplacement de capture.
Chaque femelle adulte a été équipée d’un collier Vertex Survey Globalstar (Vectronic Aerospace, Berlin, Allemagne). Ces colliers fournissent une localisation GPS toutes les 13 h ainsi qu’un signal VHF et ont été réglés pour signaler une mortalité s’ils sont stationnaires pendant 12 h. Chaque collier avait sa propre fréquence VHF unique, avec le duplicata occasionnel placé sur des individus de différentes populations entre lesquelles la dispersion était jugée improbable. Les colliers étaient également équipés d’étiquettes colorées avec des numéros uniques, permettant l’identification des individus observés sur le terrain.
L’âge de chaque femelle adulte a été estimé à partir des anneaux de croissance des cornes (Geist, 1966; Hoefs et Konig, 1984). Le sang a été obtenu par ponction veineuse jugulaire pour déterminer l’état de grossesse des femelles adultes, obtenir de l’ADN et dépister la maladie. Nous avons déterminé le statut de grossesse des femmes adultes à l’aide d’un test sérique de protéine B spécifique à la grossesse (PSPB) (Drew et coll., 2001). Les tests de grossesse des femelles adultes n’ont été effectués que l’année au cours de laquelle elles ont été capturées; des échantillons ont été envoyés à Sage Laboratories (Emmett, ID) pour effectuer des tests.
Diagnostique
Présence de M. ovipneumoniae a été détectée avec des tests de réaction en chaîne par polymérase (PCR) à l’aide d’écouvillons de bulles nasales, bronchiques et tympaniques de chaque mouflon d’Amérique capturé (Manlove et coll., 2019). Exposition précédente à M. ovipneumoniae a été déterminée en utilisant un test immuno-enzymatique compétitif (cELISA) pour détecter les anticorps dans le sérum (Ziegler et coll., 2014). Tous les tests pour M. ovipneumoniae ont été réalisées au Washington Animal Disease Diagnostic Laboratory (WADDL).
Surveillance des mouflons juvéniles
De 2016 à 2018, nous avons effectué des observations bimensuelles de toutes les femelles adultes munies d’un collier entre le 1er avril et le 31 août. L’identification des juvéniles a été déterminée par l’observation du contact physique entre les femelles adultes et les juvéniles, comme l’allaitement ou la litière ensemble. Les juvéniles sont sevrés vers l’âge de 4 mois (Festa-Bianchet, 1988); ainsi, notre période d’observation visait à couvrir la naissance jusqu’au sevrage. Nous avons localisé des femelles adultes pour l’observation à l’aide d’un récepteur de télémétrie R-1000 équipé d’une antenne directionnelle VHF RA-23K (Telonics, Inc., Mesa, AZ). Nous avons effectué des observations avec des longues-vues Kowa TSN-601 équipées d’un grossissement de 20 à 60x montées sur des trépieds. Une fois qu’une femelle adulte a été confirmée comme n’ayant pas ou plus de juvénile, c’est-à-dire deux observations consécutives où la femelle adulte a été observée sans agneau, nous avons arrêté de suivre cette femelle adulte individuelle (par exemple, Cassirer et Sinclair, 2007).
Nous avons localisé des juvéniles morts de manière opportuniste et avons prélevé des échantillons, y compris le cadavre entier, le pincement (cœur, foie et poumons), la tête, les écouvillons nasaux et auriculaires et / ou des échantillons de tissus en fonction de l’état de décomposition. Des écouvillons ont été insérés dans des flacons secs ou contenant du bouillon de soja tryptique, et d’autres échantillons ont été placés dans une glacière jusqu’à leur retour à notre base de terrain. Nous avons ensuite conservé les échantillons et les écouvillons à -20 ° C jusqu’à leur soumission au laboratoire. WADDL a réalisé une macropathologie et une histopathologie sur des échantillons de tissus pulmonaires et des tests PCR sur des écouvillons. Aditionellement, M. ovipneumoniae le typage des souches à l’aide de tests de typage de séquence multi-locus (MLST) avec quatre séquences de locus, des régions d’espacement intergéniques 16-23S, la petite sous-unité ribosomale, les gènes codant pour l’ARN polymérase B et la gyrase B, a été réalisé sur des tissus pulmonaires et des échantillons sur écouvillon provenant de deux des mortalités juvéniles récupérées (Cassirer et coll., 2017). Nous avons ensuite comparé ces types de souches à des échantillons de type souche de la métapopulation de Santa Rosa (n = 6) et des crotales (n = 2). WADDL a mené la M. ovipneumoniae typage des souches.
Échantillonnage génétique
Nous avons utilisé une combinaison des deux échantillons de sang (n = 125) et les matières fécales (n = 66) comme sources d’échantillons d’ADN. Le sang total (3 ml) fourni par ODFW et NDOW de mouflons d’Amérique capturés a été collecté dans des tubes EDTA et centrifugé à 4000 × g pendant 10 min pour séparer la couche leucocytaire. Nous avons extrait l’ADN de ce matériel à l’aide d’un kit Qiagen DNeasy Blood and Tissue (Qiagen Inc., Valencia, CA, USA). Des échantillons fécaux ont été prélevés de manière opportuniste tout en effectuant des observations de mouflons d’Amérique dans les différentes populations, et étaient généralement âgés d’une semaine ou moins, selon les estimations de la couleur, de l’odeur et de l’état de surface des granulés. Les échantillons fécaux qui étaient encore humides après le dépôt ont été séchés puis conservés à température ambiante. Les culots fécaux ont été grattés pour cibler les cellules épithéliales séchées à la surface du culot (Wehausen, Ramey et Epps, 2004), et nous avons extrait l’ADN du matériel gratté en utilisant une version modifiée du protocole Aquagenomic Stool and Soil (Multitarget Pharmaceuticals LLC, Colorado Springs, CO; voir les détails dans l’annexe S4).
Génotypage, marqueurs, identification individuelle et marqueur
Nous avons utilisé une suite de 16 marqueurs microsatellites dans trois panels (tableau S2) qui avaient précédemment été utilisés pour étudier la connectivité des populations et la variabilité génétique chez le mouflon d’Amérique (Creech et al., 2020; Creech et al., 2020; Epps, Crowhurst et Nickerson, 2018). Le génotypage a suivi les protocoles décrits dans Epps, Crowhurst et Nickerson (2018). En bref, tous les échantillons ont été exécutés dans au moins deux (pour le sang) ou trois (pour les fèces) réactions de PCR indépendantes pour générer un génotype consensus pour chaque individu à chaque locus. Pour les échantillons de sang, tout écart entre les deux répliques a entraîné une nouvelle analyse de l’échantillon dans ce panel, bien que la cohérence entre les répliques soit très élevée. Parce que l’abandon allélique peut être plus élevé dans les échantillons fécaux, pour ces échantillons, un génotype homozygote a été considéré comme vérifié si l’allèle unique a été vu dans les trois réplicats. Un génotype hétérozygote a été considéré comme vérifié si chaque allèle a été vu dans au moins deux des trois réplicats; toute autre divergence a entraîné des rediffusions. D’autres études sur le mouflon d’Amérique (p. Ex., Epps, Crowhurst et Nickerson, 2018) ont rapporté le dépistage des recaptures en utilisant aussi peu que 6 loci pour obtenir une probabilité d’identité souhaitée (PID; Waits, Luikart et Taberlet, 2001) de <0,001 et une probabilité d'identité pour les frères et sœurs à part entière de <0,05. Cependant, notre génotypage initial a démontré que nous devions génotyper les 16 loci pour atteindre ces seuils. Nous avons identifié les individus recapturés en utilisant cervus (Kalinowski, Taper et Marshall, 2007) en recherchant les individus qui correspondent aux 16 loci et en les supprimant de l’ensemble de données. Nous avons répété cette analyse en utilisant des nombres successivement réduits de locus correspondants et la présence d’un à deux mésappariements (pour tenir compte des données manquantes et des erreurs de génotypage, respectivement), jusqu’à ce que les correspondances renvoyées par le programme semblent improbables en raison de l’emplacement géographique et / ou des mésappariements. n’étaient pas explicables par un simple décrochage allélique. Enfin, nous avons utilisé vrille (Valière, 2002) pour calculer deux types de taux d’erreur dans nos génotypes: le décrochage allélique et la présence de faux allèles.
Déséquilibre de liaison, tests de Hardy-Weinberg et diversité génétique
Nous avons utilisé genepop Version 4.2 sur le Web (Rousset, 2008) pour effectuer le test de probabilité de l’équilibre de Hardy-Weinberg (HWE) pour chaque population par locus, puis pour chaque locus par population, ainsi qu’entre les populations pour chaque locus et entre les loci pour chaque population (méthode de Fisher, Fisher 1948). Nous avons ensuite utilisé genepop 4.0 Bureau pour tester le déséquilibre de liaison entre chaque paire de locus au sein de chaque population et chaque paire de locus dans toutes les populations, en appliquant une correction de Bonferroni séquentielle dans les deux cas sur tous les locus. Nous avons utilisé le package R la diversité (Keenan et coll., 2013) pour calculer les paramètres de diversité génétique de la population, y compris l’hétérozygosité attendue (HE), hétérozygotie observée (HO) et la richesse allélique (UNER). Nous avons pris en compte les tailles d’échantillon déséquilibrées parmi les populations en utilisant la raréfaction.
Données NDVI
Nous avons utilisé des données NDVI composites de 14 jours, résolution de 250 m du spectroradiomètre d’imagerie à résolution modérée (eMODIS). Nous avons utilisé des données prétraitées de 2016-2018 obtenues auprès d’Earth Explorer (https://earthexplorer.usgs.gov/), qui est géré par le Earth Resources Observation Center de la United States Geological Survey (Jenkerson, Maiersperger et Schmidt, 2010).
Nous avons utilisé les données GPS de toutes les femelles adultes munies d’un collier dans chaque population pour générer une distribution d’utilisation unique par population pour chaque année de 2016 à 2018, en utilisant le package R adehabitatHR (Calenge, 2006). Nous avons estimé les distributions d’utilisation de 95% en utilisant la méthode du noyau avec le paramètre de lissage par défaut (hréf). Nous n’avons pas utilisé la validation croisée des moindres carrés (hlscv) en raison de l’utilisation répétée d’emplacements, ce qui peut entraîner des problèmes de convergence (Kie et coll., 2010).
Nous avons ensuite extrait les données NDVI de chaque polygone de population pour chaque image composite de 14 jours et généré une statistique du 90e centile à l’aide du programme R (Équipe de base R, 2019). Les mouflons d’Amérique sont des nourrisseurs sélectifs; en tant que tel, nous supposons que la statistique NDVI du 90e percentile représente un choix de fourrage de haute qualité, tout en tenant compte du fait que le maximum de fourrage n’est pas toujours réalisable (Creech et coll., 2016). De plus, la sélection du 90e centile exclut la sélection de fausses valeurs maximales causées par une erreur de mesure. Enfin, nous avons fait la moyenne des valeurs de NDVI pour chaque composite de 14 jours pour les 3 mois avant et après le premier juvénile observé dans chaque population et avons utilisé cette variable comme mesure de la qualité du fourrage pour chaque population avant et après la parturition.
Facteurs de survie juvénile
Nous avons analysé nos données sur le devenir connu dans le programme marque (White et Burnham, 1999), pour estimer la survie des juvéniles (S) à l’aide d’un estimateur de Kaplan – Meier (Kaplan et Meier, 1958) avec entrée décalée (Pollock et coll., 1989). Premièrement, nous avons considéré trois mesures de l’exposition au niveau de la population M. ovipneumoniae dans nos modèles de survie juvénile (tableau 1). Principalement, nous avons considéré la présence de M. ovipneumoniae– juvéniles infectés dans chaque population, car le nombre limité d’adultes capturés et testés dans chaque population et le manque de tests annuels ont empêché des estimations claires des taux d’infection chez les adultes. Cependant, nous avons également examiné si la présence d’adultes infectés (PCR) ou d’adultes exposés (cELISA) dans chaque population influençait la survie des juvéniles. Deuxièmement, nous avons considéré deux mesures de la diversité génétique au niveau de la population, l’hétérozygotie attendue (HE) et la richesse allélique (UNER) (Tableau 1). Nous avons utilisé des modèles univariés des trois M. ovipneumoniae et les deux mesures de la diversité génétique comme méthodes de dépistage initial, en utilisant le critère d’information d’Akaike, corrigé pour les échantillons de petite taille (AICc) pour déterminer lequel M. ovipneumoniae et la mesure de la diversité génétique la plus fortement liée à la survie des juvéniles avant de procéder aux analyses ultérieures.
Description des variables prises en compte dans les modèles de devenir connu prédisant la survie des mouflons juvéniles (Ovis canadensis) dans les populations du sud-est de l’Oregon et du nord du Nevada.
Toutes les mesures statistiques ont été prises en compte au niveau de la population.
| Mesure | Catégorie | Type de mesure | Mesure statistique(gamme) |
|---|---|---|---|
| . (nul) | Modèle d’interception uniquement | ||
| Temps | Temporel | Variable dans le temps | |
| T | Temporel | Tendance temporelle linéaire | |
| M. ovipneumoniae statut | Les bactéries | M. ovipneumoniae statut (+/-), tel que déterminé par la présence de juvéniles infectés. | Binaire |
| M. ovipneumoniae statut | Les bactéries | M. ovipneumoniae statut (+/-) déterminé par la présence d’adultes infectés (PCR +) | Binaire |
| M. ovipneumoniae statut | Les bactéries | M. ovipneumoniae statut (+/-) déterminé par la présence d’adultes infectés (cELISA +) | Binaire |
| UNER (Richesse allélique) | Génétique | Mesure de la diversité génétique | Continu (valeurs entre 1,82 et 2,88) |
| HE (Hétérozygotie attendue) | Génétique | Mesure de la diversité génétique | Continu (valeurs entre 0 et 1) |
| Pré-NDVI (NDVI pré-parturition) | Nutrition | 90e percentile NDVI moyen avant la parturition à 3 mois | Continu (valeurs entre −0,2–1) |
| Post-NDVI (NDVI post-parturition) | Nutrition | 90e percentile 3 mois après la parturition NDVI moyen | Continu (valeurs entre −0,2–1) |
Par la suite, nous avons effectué deux analyses de la survie des juvéniles. Le premier comprenait toutes les populations étudiées (n = 13) et les mesures sélectionnées de M. ovipneumoniae presence in each population, population-level metrics of forage quality (3-month pre- and post-parturition NDVI values) and genetic diversity (expected heterozygosity, HE) (Table 1), and both additive and multiplicative effects of time. For each model that included the multiplicative effect of time and M. ovipneumoniae, we fixed survival interval 1 for the M. ovipneumoniae group, as no mortalities occurred during this period. The second analysis included all of the same variables except M. ovipneumoniae presence and was restricted to populations where M. ovipneumoniae-infected juveniles were not detected (n = 11, see Results). That second analysis was undertaken to determine whether effects of M. ovipneumoniae obscured the effect of the other covariates of interest. For both analyses, we used AICc and AICc weights (wje) to select the best-supported model. We included a null model in both model selection sets to evaluate model performance (Burnham & Anderson, 2002). We selected the model with the lowest AICc and highest wje as our best-supported model. We used evidence ratios between the top model and competitive models (those within 2 AICc units, to evaluate each model relative to the top model (Burnham & Anderson, 2002).
Post-study observations
After our final planned field season in 2018, the single PCR+ adult female in the RSP (of 21 tested) died of suspected bluetongue (Orbivirus spp.). Because prevalence of PCR+ adult females in this population was low (4.76% of tested—see Results), and no adult males tested PCR+ throughout the study, we considered it possible that no additional PCR+ individuals remained at RSP, potentially removing the source of infection for new juveniles. Therefore, we decided to conduct a single observation of juveniles in the RSP and BHP, populations in early August of 2019; we included BHP given its proximity to RSP. Juveniles in both these populations were typically 4-months old at that time; thus, that observation aligned closely with the 4-month juvenile survival we estimated during the study.
Résultats
Diagnostics
Between 2016 and 2018, 78 adult females were tested via cELISA to determine M. ovipneumoniae exposure, and 95 adult females were tested via PCR to determine active M. ovipneumoniae infections. The proportion of adult females PCR tested in each population varied from 0.10 to 0.43 (, Table S4). For the 10 adult females in the Santa Rosa metapopulation that were recaptured during our study and retested for infection via PCR, PCR status remained the same for the single positive individual in EML, and the rest were negative. None of the adult females in populations west of U.S. Route 95 (BSP, TCE, TCS, or TCW) showed evidence of M. ovipneumoniae exposure (Fig. 1, Table S3). However, all of the populations east of U.S. Route 95 (the four populations in the Santa Rosa Mountain’s metapopulation, as well as BHP, RSP, TMP, and UOP (Fig. 1)), except TFK, included adult females with evidence of exposure to M. ovipneumoniae (Table S3). We were unable to get a sample from the single individual adult female captured in TFK. The proportion of M. ovipneumoniae exposed adult females in those eight populations varied between 0.60 in CAL and EML to 1.00 in TMP (; Table S4). Only two adult females tested PCR+ to M. ovipneumoniae infection across all 13 populations (Table S3): one in EML on two occasions (2017 and 2018), and one in RSP in 2016. Additionally, one adult female in BHP yielded an indeterminate PCR test result, meaning M. ovipneumoniae detection could not be determined (Besser et al., 2019).
Linkage disequilibrium, Hardy-Weinberg tests, and genetic diversity
After removing the fecal samples from recaptured individuals (n = 28), the genetic data set contained 191 individuals at 16 microsatellite loci, representing 10 to 26 () individuals per population (Table S5). The mean rate of false allele occurrence per locus was 0.001 and the mean allelic dropout rate across all loci was 0.008 (range = 0.000–0.017). No locus was determined to be out of Hardy-Weinberg equilibrium by either test employed (Tables S6 & S7). Evaluating each locus pair by population showed no evidence of linkage disequilibrium (pcritical = 0.000035 for α = 0.05); evaluating linkage for each pair of loci across populations using Fisher’s test indicated that BL4 and HH62 were in disequilibrium (p = 0.00039; pcritical = 0.00042 for α = 0.05). However, those loci have not appeared to be in disequilibrium in other bighorn sheep studies (e.g., Epps, Crowhurst & Nickerson, 2018), suggesting that this relationship may have been an artifact. Thus, and because linkage disequilibrium is more likely to bias estimates of genetic structure rather than estimates of genetic diversity as employed in this study, we retained all 16 loci in our analyses. HE across the 13 study populations varied from 0.26 in BSP to 0.48 in EML (; Table S5) and UNER varied from 1.82 in BSP to 2.88 in SAW (; Table S5).
NDVI data
Pre- and post-parturition NDVI varied spatially and temporally (Table S8). BSP had consistently low pre-parturition NDVI, whilst EML and TMP had consistently high pre-parturition NDVI (Table S8). Post-parturition NDVI was lowest in BHP in 2018, was consistently low in RSP but was consistently high in TCE and EML (Table S8). Pre- and post-parturition NDVI values were not correlated (Table S9).
Pregnancy rates and observation of juvenile bighorn sheep
Seventy-six of the 82 (93%), pregnancy tests conducted across all 13 populations between 2016 and 2018 were positive (Table S10). Six of the 82 tests conducted were on recaptured adult females from EML (n = 3) and SAW (n = 3), all of which were positive. All populations had 100% pregnancy rates, except for TCE (67%; n = 8∕12), TCS (50%; n = 1∕2), and TFK (0%; n = 0∕1) in 2016. The single collared adult female in TFK did not yield a positive pregnancy test result in January, but was later observed in early June with a juvenile, suggesting that the test was administered too early to detect a positive pregnancy result. Sixty-five of those 82 (79%) pregnant adult females survived to parturition; of those 59 (91%) were observed with juveniles. Population pregnancy rates were not correlated with genetic diversity (Pearson pairwise correlation; HE, r = 0.24, p = 0.272; UNER, r = 0.14, p = 0.529).
We observed 121 juveniles with radio-collared adult females between 2016 and 2018; 78% of radio-collared adult females were observed with juveniles. Populations with the lowest proportion of radio-collared adult females with juveniles were BSP (2016—0.00; 2017—0.67; 2018—0.33), and TCE (2016—0.64; 2017—0.64; 2018—0.17), although sample sizes were small in BSP (n = 3). A set of twins was observed with a radio-collared adult female in both the BHP and MCK populations in 2018.
The observation rate of collared adult females with juveniles varied from 50 to 100%, with a mean observation rate of 94% across all semi-monthly sampling periods and study populations (Table S11). Throughout the study, we collected samples suitable for M. ovipneumoniae testing from 17 juvenile mortalities (Table S12). All juvenile mortalities (n = 15) recovered from the BHP (n = 1) and RSP (n = 14) populations tested positive for M. ovipneumoniae. None of the other juvenile mortalities from BSP (n = 1) or TCS (n = 1) tested positive for M. ovipneumoniae. Le M. ovipneumoniae strain-type of the samples collected from the BHP and RSP juveniles matched the strain-type (NV_BHS_SantaRosas_2651_2014_4; Kamath et al., 2019) of all the other previously strain-typed bighorn sheep at the available sequences from the Santa Rosa meta-population and Rattlesnake population (Table S13).
Drivers of juvenile survival
Preliminary analyses with univariate models of the relationship between juvenile survival and different population-level measures of M. ovipneumoniae revealed that the presence of M. ovipneumoniae-infected juveniles in the population was a much stronger predictor of juvenile survival than the presence of infected (PCR+) or exposed (ELISA+) adults (Table S14). Preliminary analyses with univariate models of juvenile survival as a function of genetic diversity demonstrated that HE was a stronger predictor of juvenile survival than UNER (Table S14). Therefore, in our subsequent multivariate models of juvenile survival, we used the presence of M. ovipneumoniae-infected juveniles in each population as our measure of M. ovipneumoniae presence, and HE as our measure of genetic diversity. In those multivariate analyses, we identified two competing models predicting survival probability of juveniles across all populations (Table 2). Both models contained only two predictors: (1) whether M. ovipneumoniae-infected juveniles were detected, and (2) time. The model containing time as a multiplicative effect with M. ovipneumoniae had 2.36 times more support than the competing model that treated time as an additive effect (Table 2), meaning that the temporal pattern of juvenile mortality differed in populations where M. ovipneumoniae-infected juveniles were present. Neither forage quality (pre- and post-parturition NDVI), nor genetic diversity (HE) predicted survival probability of juveniles (Table 2).
Model selection results for known fate models predicting cumulative 4-month survival of juvenile bighorn sheep (Ovis canadensis) in southeastern Oregon and northern Nevada for the period 2016–2018.
For analysis 1, we included a binary group variable for populations where Mycoplasma ovipneumoniae was (n = 2) and was not detected (n = 11). We modeled time as a constant (.), time-varying (time), linear (T), and random effect. Covariates modeled include expected heterozygosity (HE), and pre- and post-parturition NDVI. For analysis 2, we only modeled populations (n = 11) with no observed M. ovipneumoniae mortalities. We modeled time as a constant (.), time-varying (time), and linear (T) effect. We included the same covariates in data set 2 that we used in data set 1.
| Analyse | Model | K | AICc | ΔAICc | wje | ML |
|---|---|---|---|---|---|---|
| 1 | M. ovipneumoniae× time | 15* | 366.51 | 0.00 | 0.30 | 1.00 |
| M. ovipneumoniae + time | 9 | 368.23 | 1.71 | 0.13 | 0.42 | |
| M. ovipneumoniae× time + HE | 16* | 368.58 | 2.06 | 0.11 | 0.36 | |
| M. ovipneumoniae× time + post-NDVI | 16* | 368.63 | 2.12 | 0.10 | 0.35 | |
| M. ovipneumoniae× time + pre-NDVI | 16* | 368.64 | 2.12 | 0.10 | 0.35 | |
| M. ovipneumoniae + time + HE | dix | 370.02 | 3.51 | 0.05 | 0.17 | |
| M. ovipneumoniae + time + pre-NDVI | dix | 370.23 | 3.72 | 0.05 | 0.16 | |
| M. ovipneumoniae + time + post-NDVI | dix | 370.31 | 3.79 | 0.04 | 0.15 | |
| M. ovipneumoniae× time + HE + post-NDVI | 17* | 370.66 | 4.15 | 0.04 | 0.13 | |
| M. ovipneumoniae× time + HE + pre-NDVI | 17* | 370.69 | 4.18 | 0.04 | 0.12 | |
| M. ovipneumoniae× time + pre-NDVI + post-NDVI | 17* | 370.77 | 4.25 | 0.04 | 0.12 | |
| M. ovipneumoniae× time + HE + pre-NDVI + post-NDVI | 18* | 372.80 | 6.29 | 0.01 | 0.04 | |
| Time + post-NDVI | 9 | 383.02 | 16.51 | 0.00 | 0.00 | |
| Time + HE | 9 | 387.94 | 21.42 | 0.00 | 0.00 | |
| Time | 8 | 392.21 | 25.70 | 0.00 | 0.00 | |
| Time + pre-NDVI | 9 | 394.22 | 27.71 | 0.00 | 0.00 | |
| M. ovipneumoniae | 2 | 400.46 | 33.94 | 0.00 | 0.00 | |
| post-NDVI | 2 | 412.59 | 46.08 | 0.00 | 0.00 | |
| HE | 2 | 417.59 | 51.07 | 0.00 | 0.00 | |
| T | 2 | 418.76 | 52.24 | 0.00 | 0.00 | |
| . (null) | 1 | 422.68 | 56.17 | 0.00 | 0.00 | |
| pre-NDVI | 2 | 424.41 | 57.90 | 0.00 | 0.00 | |
| 2 | Time | 8 | 210.79 | 0.00 | 0.23 | 1.00 |
| . (null) | 1 | 211.84 | 1.05 | 0.14 | 0.59 | |
| Time + pre-NDVI | 9 | 212.52 | 1.73 | 0.10 | 0.42 | |
| Time + HE | 9 | 212.70 | 1.91 | 0.09 | 0.38 | |
| Time + post-NDVI | 9 | 212.78 | 1.99 | 0.09 | 0.37 | |
| T | 2 | 213.27 | 2.48 | 0.07 | 0.29 | |
| post-NDVI | 2 | 213.60 | 2.82 | 0.06 | 0.24 | |
| pre-NDVI | 2 | 213.67 | 2.88 | 0.05 | 0.24 | |
| HE | 2 | 213.69 | 2.90 | 0.05 | 0.23 | |
| Time + HE + pre-NDVI | dix | 214.19 | 3.41 | 0.04 | 0.18 | |
| Time + pre-NDVI + post-NDVI | dix | 214.41 | 3.62 | 0.04 | 0.16 | |
| Time + HE + post-NDVI | dix | 214.78 | 4.00 | 0.03 | 0.14 | |
| Time + HE + pre-NDVI + post-NDVI | 11 | 216.25 | 5.46 | 0.02 | 0.07 |
The odds of a juveniles surviving to 4-months of age in our study populations where M. ovipneumoniae-infected juveniles were detected were 8.00 (95% CI [−4.00–255.78]) times less likely than juveniles in populations where we did not detect M. ovipneumoniae-infected juveniles (Table 3). The derived probability of survival for the entire 4-month annual study period for juveniles in populations where M. ovipneumoniae-infected juveniles were detected was 0.02 (95% CI [0.00–0.13]) compared to 0.44 (95% CI [0.29–0.59]) for juveniles in other populations (Fig. 2B).
The outputs from the top models in analysis 1 (M. ovipneumoniae× time) and analysis 2 (null) predicting survival of juvenile bighorn sheep (Ovis canadensis) in southeastern Oregon and northern Nevada.
Analysis 1 includes all study populations, while analysis 2 only includes populations (n = 11) where Mycoplasma ovipneumoniae was not detected.
| Analyse | Covariate | Effect on survival | Odds-ratio | Estimate | SE | 95% CI | |
|---|---|---|---|---|---|---|---|
| Lower | Upper | ||||||
| 1 | Intercept | 2.08 | 1.06 | 0.00 | 4.16 | ||
| M. ovipneumoniae | ↓ | 8.00 | −2.08 | 1.77 | −5.54 | 1.39 | |
| Time 1 | ↑ | 9.50 | 2.25 | 1.46 | −0.61 | 5.12 | |
| Time 2 | ↑ | 1.00 | 0.00 | 1.13 | −2.22 | 2.22 | |
| Time 3 | ↓ | 1.89 | −0.64 | 1.12 | −2.84 | 1.56 | |
| Time 4 | ↓ | 1.05 | −0.05 | 1.16 | −2.33 | 2.23 | |
| Time 5 | ↑ | 1.83 | 0.61 | 1.22 | −1.78 | 2.99 | |
| Time 6 | ↑ | 2.44 | 0.89 | 1.28 | −1.63 | 3.41 | |
| Time 7 | ↓ | 1.09 | −0.09 | 1.23 | −2.49 | 2.32 | |
| M. ovipneumoniae× time 1* | ↑ | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| M. ovipneumoniae× time 2 | ↑ | 4.37 | 1.48 | 1.85 | −2.16 | 5.11 | |
| M. ovipneumoniae× time 3 | ↑ | 4.36 | 1.47 | 1.85 | −2.15 | 5.09 | |
| M. ovipneumoniae× time 4 | ↓ | 1.33 | −0.29 | 1.88 | −3.96 | 3.39 | |
| M. ovipneumoniae× time 5 | ↓ | 2.75 | −1.01 | 1.97 | −4.88 | 2.86 | |
| M. ovipneumoniae× time 6 | ↓ | 1.63 | −0.49 | 2.12 | −4.64 | 3.66 | |
| M. ovipneumoniae× time 7 | ↑ | 2.18 | 0.78 | 2.24 | −3.60 | 5.16 | |
| 2 | Intercept | 2.08 | 1.06 | 0.00 | 4.16 | ||
| Time 1 | ↑ | 9.50 | 2.25 | 1.46 | −0.61 | 5.12 | |
| Time 2 | ↓ | 1.00 | 0.00 | 1.13 | −2.22 | 2.22 | |
| Time 3 | ↓ | 1.89 | −0.64 | 1.12 | −2.84 | 1.56 | |
| Time 4 | ↓ | 1.05 | −0.05 | 1.16 | −2.33 | 2.23 | |
| Time 5 | ↑ | 1.83 | 0.61 | 1.22 | −1.78 | 2.99 | |
| Time 6 | ↑ | 2.44 | 0.89 | 1.28 | −1.63 | 3.41 | |
| Time 7 | ↓ | 1.09 | −0.09 | 1.23 | −2.49 | 2.32 | |
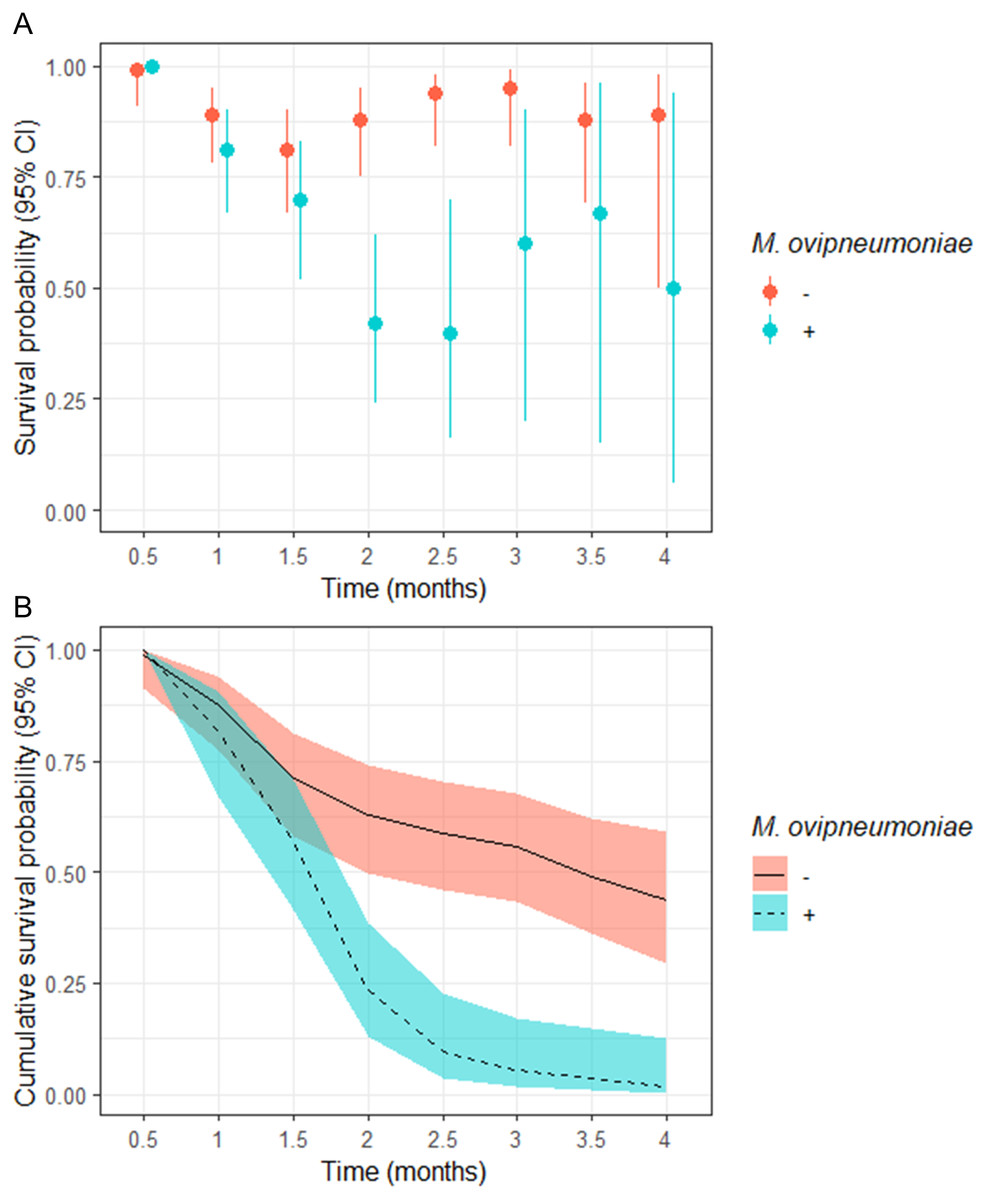
Figure 2: (A) Semi-monthly survival probabilities and (B) 4-month cumulative survival probabilities of juvenile bighorn sheep (Ovis canadensis) in populations across southeastern Oregon and northern Nevada.
(A) Semi-monthly survival probabilities and (B) 4-month cumulative survival probabilities of juvenile bighorn sheep (Ovis canadensis) estimated using a Kaplan–Meier estimator with staggered entry and known-fate data collected in 13 populations in southeastern Oregon and northern Nevada. Survival probabilities and 95% confidence intervals are for populations where Mycoplasma ovipneumoniae was and was not detected.
We observed no significant difference in semi-monthly juvenile survival probability between populations where M. ovipneumoniae-infected juveniles were not detected in observations at 0.5, 1, and 1.5-months post-parturition (Fig. 2A). However, semi-monthly survival probability in populations where M. ovipneumoniae-infected juveniles were present were significantly lower 2-months (S exposed = 0.42, 95% CI [0.24–0.62]; S unexposed = 0.88, 95% CI [0.75–0.95]) and 2.5-months (S exposed = 0.40, 95% CI [0.16–0.70]; S unexposed = 0.94, 95% CI [0.82–0.98]) post-parturition (Fig. 2A). At 3, 3.5, and 4-months post-parturition, estimates in semi-monthly survival probability again did not differ between populations where M. ovipneumoniae-infected juveniles were or were not detected (Fig. 2A). The increase in uncertainty in our estimates is tied to decreasing sample size as individuals died.
Although no direct effect of either population genetic diversity or post-parturition NDVI was supported based on our model selection approach, both variables were highly correlated with the presence of M. ovipneumoniae (HE = 0.58; post-NDVI = −0.66, Table S9). In fact, the two populations where M. ovipneumoniae-infected juveniles were present, BHP and RSP, had relatively high genetic diversity (HE: BHP = 0.43; RSP = 0.45) compared to other study populations (Table S5). Similarly, RSP had low post-parturition NDVI values relative to the other populations in 2017 and 2018, while the same was the case for BHP in 2018 (Table S8).
In the second analysis, which excluded the two populations where M. ovipneumoniae-infected juveniles were present, competing models included both indices of forage quality and population genetic diversity as predictors of juvenile survival (Table 2), as well as a time-varying effect. However, the 95% confidence intervals for the parameter estimates of the nutritional and population genetic diversity model both overlapped zero, and the null model with no predictors was 2nd ranked. The top competing model, a time-varying model, had 1.69 times more support than the next best model, the null model (Table 2).
Post-study observations
In early August of 2019, following the death of the single adult female in RSP that tested PCR+ for M. ovipneumoniae, we located the two remaining collared individuals in the BHP (linked to RSP by adult male and female movements, data not shown). Neither the collared adult females nor any of the eight other adult females we observed in this population had juveniles (0% juvenile survival). We then located 14 of the RSP collared adult females, of which 11 had juveniles (∼79% juvenile survival). We located approximately 35 adult females total in the RSP population and observed approximately 27 juveniles, which equates to an approximate 4-month survival rate of ∼77%.
Discussion
We found that M. ovipneumoniae had a strong negative effect on juvenile survival in populations where infected juveniles were detected. Juveniles in those populations had a 4-month survival probability more than 20 times lower than juveniles in populations where we did not detect infected juveniles. Indeed, in one affected population (RSP), only two juveniles out of 40 over three years survived. Presence of adult bighorn sheep with evidence of past exposure to M. ovipneumoniae, and in one population the presence of an adult infected at time of capture, were not strong predictors of juvenile survival. Moreover, juvenile survival was not detectably influenced by forage quality or population genetic diversity, even when excluding populations where M. ovipneumoniae-related juvenile mortality was observed. Strikingly, the death of a single infected adult female in a heavily impacted population appeared to drive an immediate recovery of juvenile survival in that population in the subsequent year, underscoring the potentially pivotal role of chronically infected adult females in outbreaks of this disease.
In our study, four-month juvenile survival probability in populations where infected juveniles were present (0.02; 95% CI [0.00–0.13]) were comparable to some of the lowest observed juvenile survival probabilities reported in other studies where pneumonia epizootics were present. In Rocky Mountain bighorn sheep in the Hells Canyon system of Idaho, Oregon, and Washington, survival to weaning at 4–5 months of age had a median value of 0.10, but ranged from <0.01–0.69, with peak mortality occurring between 1.5 and 2.5 months of age (Cassirer et al., 2013). In the Black Hills of South Dakota, Smith et al. (2014) reported 52-week survival to be only 0.02 (95% CI [0.01–0.07]), with the majority of juveniles (∼75%) dying within the first 2 months after parturition. We observed similar temporal patterns to those studies, with most mortality related to disease in our study occurring between 1.5 and 3-months after parturition. In the Mojave Desert of California, in a 3-year study of desert bighorn sheep following an outbreak of M. ovipneumoniae, the proportion of juveniles surviving varied from 0.00 to 0.92 in populations where at least five juveniles were monitored, and survival in most populations improved in later years (Dekelaita et al., 2020). Unlike Cassirer et al. (2013) et Smith et al. (2014), we did not address the effect of predators on juvenile survival. However, it should be noted that even if cause-specific mortality were to be assigned to a predator, the ultimate cause of mortality might still be M. ovipneumoniae. Although M. ovipneumoniae is considered the primary agent of pneumonia, other diseases, and the potential for co-infection effects may be important, but we did not assess them. Periods of low juvenile survival due to pneumonia can result in demographic consequences such as declining population growth rates (Manlove et al., 2016) and even skewed population structures (Festa-Bianchet, Gaillard & Côté, 2003).
Our study adds to the evidence that dynamics of respiratory pneumonia in bighorn sheep may be strongly influenced by the presence of only a small number of infected adults. We detected only one infected female adult bighorn sheep across two populations where M. ovipneumoniae-infected juvenile bighorn sheep were observed. However, the presence of infected adults at capture was not the strongest predictor of juvenile survival. For instance, in EML, an infected female adult was present in 2017; she was still infected when retested in 2018. Despite that, one out of four juveniles born in 2017 and five of six juveniles born in 2018 survived. It is unclear why the presence of this apparently chronically-infected female did not lead to juvenile mortality in this case, although Plowright et al. (2017) noted that juvenile survival was not directly related to the mother’s infection status in the study. The lack of juvenile die-off could be explained by other factors, e.g., the stochastic nature of exposure and the possibility of co-infection. We attempted to capture other factors that may play a role, e.g., measures of forage quality and population genetic diversity, but we did not detect any direct effect of those factors. Additionally, we were unable to assess the interaction of these variables with M. ovipneumoniae infection because our measures of those factors were at the population level. Conversely, juvenile survival in RSP was extremely poor from 2016–2018, where a single infected adult female was detected in 2016. Although we did not retest her and therefore do not know whether she was chronically infected, after her death in late 2018, juvenile survival rebounded sharply in 2019. In a satellite population discovered in the course of this study, BHP, one female out of three captured in 2018 yielded an indeterminate PCR result. We surmise that additional captures likely would have detected infected adults given that infected juveniles were detected in 2018 and no surviving juveniles were detected in either 2018 or 2019. Experimental removals of chronically-infected adult females in other bighorn sheep studies (e.g., Garwood et al., 2020) demonstrate that chronically-infected adult females can strongly influence juvenile survival when prevalence is low.
We did not detect influences of population genetic diversity on juvenile survival, although we observed low to very low population genetic diversity throughout our study area, with HE ranging from 0.26–0.48 (Table S5). In contrast, HE values derived for 11 native desert bighorn sheep populations in the Mojave Desert of California, using 15 of the 16 microsatellite loci we employed, ranged from 0.50–0.69 (Epps, Crowhurst & Nickerson, 2018). However, our study design did not account for possible interactions between these variables and presence of M. ovipneumoniae-infected juveniles, because M. ovipneumoniae-related juvenile mortalities were confirmed in only two of 13 populations. Our estimates of genetic diversity were also only derived from 16 microsatellite loci, most putatively neutral; estimates based on more markers across the genome or additional genes associated with immune function could have been more informative. Interestingly, the populations where M. ovipneumoniae-related juvenile mortality was confirmed had relatively high genetic diversity (Table S5). In fact, the higher genetic diversity observed at RSP likely resulted from gene flow from males dispersing from the Santa Rosa metapopulation (NDOW, unpublished data). Such movements also appeared to have spread this strain of M. ovipneumoniae from the Santa Rosa metapopulation to BHP and RSP; indeed, only one strain is known to occur throughout the populations where we detected infected individuals (Table S13). Genetic diversity in populations to the west of U.S. Route 95, in particular, was strikingly low (HE was 0.26–0.33) due to founder effects from translocation history (see above). Genetic diversity observed by Olson, Whittaker & Rhodes (2013) for the source population, Hart Mountain, was noticeably higher (HE = 0.42). We note that the pregnancy rate in one of those populations (TCE) likewise was strikingly low (67%, n = 8∕12) in 2016. Although the adult females that we had the opportunity to test in 2017 and 2018 (n = 3 total) were pregnant, the numbers of adult females observed with juveniles in 2017 and 2018 were equal to or lower than 2016. Yet, those populations were apparently free of M. ovipneumoniae during our study. Such low pregnancy rates suggest that inbreeding or low genetic diversity could influence performance of those populations (Olson, Whittaker & Rhodes, 2012), although we cannot rule out other factors. These contrasts highlight the challenges of maintaining genetic diversity while limiting spread of disease, particularly in reintroduced systems of populations where compounding founder effects have resulted in some of the lowest reported values for genetic diversity in wild bighorn sheep populations.
Forage quality as assessed by NDVI values pre- and post-parturition likewise did not appear to influence juvenile survival consistently across the study. Again, our ability to evaluate how variation in forage quality influenced survival when M. ovipneumoniae was present was limited. We note, however, that of the three populations where M. ovipneumoniae-infected adult females or juveniles were detected, juvenile survival was lowest where forage quality was lowest (BHP and RSP, Table S8, average juvenile survival = 0 and 0.07, respectively). In contrast, EML had the highest or second-highest pre- and post- parturition NDVI values (Table S8) and higher juvenile survival (; range = 0.25–0.83). Taken together, these anecdotal patterns suggest that the link between forage quality and M. ovipneumoniae dynamics within infected populations bears further investigation.
Our study demonstrated that reintroduced populations of “California”-managed bighorn sheep in southeastern Oregon and northern Nevada have been strongly influenced by respiratory disease, but not all populations are currently affected. Where infected juveniles were present, rates of juvenile mortality that we observed would lead to a dramatic decline in populations if those patterns persisted, as observed by Manlove et al. (2016). However, the rapid improvement in juvenile survival in one badly-affected population following the death of the single adult female known to be infected with M. ovipneumoniae at the start of the study suggests that identification and removal of chronically-infected adult females could be a successful management strategy in this system, as tested for a Rocky Mountain bighorn sheep population by Garwood et al. (2020). The very low estimates of genetic diversity we observed in some populations suggest that actions to manage for improved genetic diversity may be warranted, although natural movements between several study populations simultaneously resulted in increased genetic diversity and spread of M. ovipneumoniae. Thus, any genetic management strategy employed would have to mitigate the potential impact on spread of disease.
Conclusions
We found that populations with M. ovipneumoniae-infected juveniles had significantly lower juvenile survival, while no direct effect of either genetic diversity or pre- and post-parturition forage quality on juvenile survival was detected. Presence of M. ovipneumoniae-infected or exposed adults was less predictive of juvenile survival. Our findings add to the body of evidence that M. ovipneumoniae can have deleterious effects on juvenile survival across the various bighorn sheep lineages and the different ecosystems they inhabit. After the mortality of the only PCR+ adult female that we detected in one of the populations most affected by respiratory disease during the first three years of our study, we found increased juvenile survival and detected no infected juveniles in the following year. This natural experiment suggests that in populations where prevalence of PCR+ adults is low, removal of infected adults or adults identified as chronic carriers of M. ovipneumoniae. However, our ability to detect those influences may have been limited by small number of populations where respiratory disease was observed and high correlations between genetic diversity and presence of disease resulting from translocation history and patterns of connectivity in this system. Future studies should evaluate the interaction of genetic diversity and disease in systems with larger numbers of populations affected by respiratory disease.
Supplemental Information
Raw data and outputs used in figures
Raw data utilized in Program Mark to generate results, and associated figures, and tables.
Source de l’article